Cette immunité sociale résulte de la coopération des individus de la colonie dans le but de lutter contre le risque accru de transmission de maladies qui découle de la vie en société. Les défenses collectives peuvent être à la fois prophylactiques et activées à la demande. Il s’agit parfois d’adaptations comportementales, physiologiques et organisationnelles de la colonie qui empêchent l’entrée, l’établissement et la propagation des vecteurs de maladie.
Si la vie en société présente de nombreux avantages, en particulier en ce qui concerne l’efficacité des soins au couvain, la recherche de nourriture et la défense de la colonie, ces avantages peuvent également présenter quelques inconvénients. Les maladies infectieuses se propagent plus facilement entre les membres d’un groupe du fait de la densité élevé d’individus et des contacts sociaux fréquents. Les sociétés animales complexes ont développé des stratégies pour pallier ces risques.
Ces défenses sociales reposent sur une action collective et sur des comportements altruistes d’individus infectés qui profitent à la colonie. Ces défenses dépendent de la coopération des membres du groupe social pour éviter, contrôler ou éliminer les infections et leurs sources.
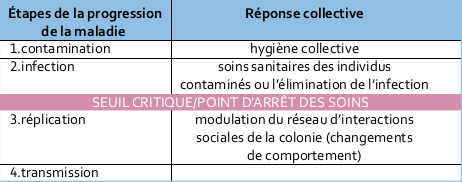
La progression de la maladie et les étapes de l’immunité sociale
L’immunité sociale agit pour interférer à chaque étape de la progression de la maladie. Pour protéger la reine et la colonie des maladies, des mécanismes de défense coopératifs distincts sont employés à chaque étape. En cas d’échec, la maladie passe à l’étape suivante et l’agent pathogène infecte progressivement les individus de la colonie, se réplique et se transmet même potentiellement à d’autres colonies. La colonie commence par se protéger en protégeant ses membres individuels par des comportements de nettoyage
par exemple. La colonie se prémunit aussi contre la transmission en prenant soin du nid via les propriétés antimicrobiennes de la propolis. D’autres défenses sont activées à la demande, par exemple la fièvre sociale où de nombreuses abeilles augmentent simultanément leur température corporelle. Concernant l’élimination de l’infection, on sait par exemple que les abeilles peuvent détecter le couvain infecté à travers l’opercule. Certaines abeilles désoperculent et retirent le couvain infecté pour le déposer au sol
loin de la colonie.
Une fois le seuil critique atteint, les changements de comportement apparaissent qui peuvent aller jusqu’au sacrifice des individus infectés. Les abeilles immunodéprimées soit se bannissent de manière altruiste soit sont expulsées par des congénères qui ont identifié le problème à l’aide d’indices olfactifs.
Ainsi, des gardiennes spécialisées qui contrôlent l’entrée du nid peuvent attaquer et exclure les ouvrières infectées.
La température du nid
Comportement hygiénique, élimination du couvain malade, collecte et dépôt de propolis sur les surfaces intérieures du nid sont des comportements désormais bien connus des apiculteurs. La fièvre sociale est une autre forme d’immunité collective moins connue.
Les abeilles mellifères augmentent la température du nid en réponse, par exemple, à Ascosphaera apis, agent pathogène du couvain sensible à la température, responsable de la maladie du couvain sacciforme.
L’augmentation de la température du nid limiterait la propagation de l’infection au couvain non infesté. Certaines études indiquent que les ouvrières exposées à un agent pathogène du couvain élèvent la température du nid à couvain et initient ainsi une réponse immunitaire.
La ventilation simultanée des ailes de nombreuses ouvrières à l’intérieur de la colonie augmente la température de la ruche de plusieurs degrés. Ce mode de défense est particulièrement employé contre les infections de la colonie par des spores fongiques.
On connait bien l’importance du maintien de la température du couvain à une fourchette de 33 à 36°C. On sait déjà que des températures sous-optimales peuvent entraîner des anomalies à l’émergence et se traduisent par divers problèmes chez les abeilles adultes en lien avec l’apprentissage, la mémoire, la morphologie, la spécialisation des tâches, les performances, la résistance au stress, la physiologie de la reproduction, la longévité, etc. L’importance d’un bon maintien de la thermorégulation du nid et la nécessité de favoriser ce besoin vital pour les abeilles doit être bien comprise par les apiculteurs.
Une température du couvain inférieure à la normale et une humidité relative élevée peuvent provoquer des maladies et accélérer la mortalité chez un pourcentage important de larves exposées à l’agent pathogène.
L’amorçage immunitaire transgénérationnel
L’immunité sociale comprend aussi le transfert oral de composés immunologiques entre les ouvrières. La gelée royale contient de nombreux composés antimicrobiens. Une forme connexe d’immunité sociale appelé amorçage immunitaire transgénérationnel, permet aux reines de transférer des fragments d’agents pathogènes dans leurs œufs en développement où ils sont reconnus par le système immunitaire de l’embryon et induisent une plus grande résistance aux agents pathogènes chez la nouvelle progéniture.
Ces fragments pathogènes sont transportés par la vitellogénine (protéine produite par les nourrices pour synthétiser la gelée royale). La gelée royale peut ainsi servir de véhicule pour transporter des fragments d’agents pathogènes. La consommation de cellules pathogènes induirait des niveaux plus élevés d’un peptide antimicrobien présent dans la gelée royale appelé « défensine-1 ».
L’amorçage immunitaire transgénérationnel peut fonctionner à l’échelle de la colonie. En plus d’un transfert vertical de la reine à sa progéniture, les ouvrières peuvent être responsables d’une voie plus complexe allant des nourrices à la reine et aux futures larves, ainsi qu’une transmission horizontale des ouvrières aux larves. L’avantage serait que les larves sont inoculées avec un répertoire plus large d’éliciteurs immunitaires (substances capables de stimuler les mécanismes de défense naturelle) accumulés à partir de l’expérience immunologique collective des ouvrières qui collectent des ressources à
l’extérieur du nid. Cela rendrait les larves plus résistantes aux agents pathogènes qu’elles sont susceptibles de rencontrer lorsqu’elles sortiront à leur tour hors du nid.
La diversité génétique
Il reste à citer l’importance de la diversité génétique dans le contexte de l’immunité sociale. La diversité génétique au sein de la colonie façonne les microbiotes associés aux abeilles mellifères d’une manière telle que les répercussions sont importantes sur le fonctionnement et la santé de la colonie. La diversité génétique contribue ainsi de manière prophylactique à la réduction des risques sanitaires.
La résistance de la colonie est augmentée par le nombre de partenaires d’accouplement de la reine. Les avantages d’une diversité génétique accrue résulterait de la recombinaison génomique, qui se produit à des taux élevés chez plusieurs espèces eusociales. Les colonies hétérogènes seraient susceptibles
de subir moins de dommages que les colonies homogènes, y compris dans une certaine mesure en ce qui concerne la défense contre les parasites même si de nombreux paramètres entre en jeu à ce niveau comme la fréquence des infections et le taux d’infestation.
Comme toutes les sociétés d’insectes sociaux, les abeilles mellifères ont développé des systèmes immunitaires sociaux très complexes qui forment des barrières fonctionnelles et qui proposent des réponses prophylactiques et des réponses activées en fonction des besoins. Pour la colonie, il s’agit d’adapter son comportement ou son organisation pour empêcher l’entrée, la propagation et l’établissement à long terme de problèmes allant de la propagation de parasites, d’infections virales ou bactériennes ou encore d’attaques de prédateurs.
Références :
Cini, A., Bordoni, A., Cappa, F., Petrocelli, I., Pitzalis, M., Iovinella, I., ... & Cervo, R. (2020). Increased immunocompetence and network centrality of allogroomer workers suggest a link between individual and social immunity in honeybees. Scientific reports, 10(1), 1-12.
Conroy, T. E., & Holman, L. (2022). Social immunity in the honey bee : do immune-challenged workers enter enforced or self-imposed exile ? Behavioral Ecology and Sociobiology, 76(2), 1-9.
Cremer, S. (2019). Social immunity in insects. Current Biology, 29(11), R458-R463.
Drum, N.H., and Rothenbuhler, W.C.(1985). Differences in non- stinging aggressive responses of worker honeybees to diseased and healthy bees in May and July. J. Apicult. Res. 24, 184–187.
Evans JD et al. 2006 Immune pathways and defence mechanisms in honey bees Apis mellifera. Insect Mol. Biol. 15, 645 – 656. (doi:10.1111/j.1365-2583. 2006.00682.x)
Goblirsch, M., Warner, J. F., Sommerfeldt, B. A., & Spivak, M. (2020). Social fever or general immune response ? Revisiting an example of social immunity in honey bees. Insects, 11(8), 528.
Harwood, G., Salmela, H., Freitak, D., & Amdam, G. (2021). Social immunity in honey bees : royal jelly as a vehicle in transferring bacterial pathogen fragments between nestmates. Journal of Experimental Biology, 224(7), jeb231076.
Jones, B., Shipley, E., & Arnold, K. E.(2018). Social immunity in honeybees—Density dependence, diet, and body mass trade‐offs. Ecology and evolution, 8(10), 4852-4859.
Liu, L., Zhao, X. Y., Tang, Q. B., Lei, C. L., & Huang, Q. Y.(2019). The mechanisms of social immunity against fungal infections in eusocial insects. Toxins, 11(5), 244.
Mattila, H. R., Rios, D., Walker-Sperling, V. E., Roeselers, G., & Newton, I. L.(2012). Characterization of the active microbiotas associated with honey bees reveals healthier and broader communities when colonies are genetically diverse. PLoS One, 7(3), e32962.
Meunier, J. (2015). Social immunity and the evolution of group living in insects. Philosophical Transactions of the Royal Society B : Biological Sciences, 370(1669), 20140102.
Simone-Finstrom, M., & Spivak, M. (2010). Propolis and bee health : the natural history and significance of resin use by honey bees. Apidologie, 41(3), 295-311.
Starks PT, Blackie CA, Seeley TD. 2000 Fever in honeybee colonies. Naturwissenschaften 87, 229– 231. (doi:10.1007/s001140050709)
Starks, P.T., Blackie, C.A., and Seeley, T.D.(2000). Fever in honey bee colonies. Naturwissenschaften 87, 229–231.
Van Baalen M, Beekman M. 2006 The costs and benefits of genetic heterogeneity in resistance against parasites in social insects. Am. Nat. 167, 568 – 577. (doi:10.1086/501169)
Waddington, K.D., and Rothenbuhler, W.C.(1976). Behaviour associated with hairless-black syndrome of adult honeybees. J. Apicult. Res. 15, 35–41.