Les facteurs saisonniers
D’une manière générale, la population de mâles dans une colonie augmente au printemps et atteint un sommet à la fin du printemps ou au début de l’été. Le pic de la produc tion de mâles coïncide avec le pic de production d’ouvrières, juste avant la période d’essaimage c’est-à-dire le temps de la reproduction de la colo nie. Un grand nombre de mâles mûrs à cette période augmente le succès reproductif. Après cette période que l’on peut estimer correspondre à mai-juin-juillet, la population de mâles diminue progressivement pendant l’été et en automne pour généralement être à zéro en hiver. Il y a cependant des exceptions.
Cette tendance générale est influencée par la taille de la colonie, la disponibilité de la nourriture et également le microclimat de la colonie.
La taille de la colonie
Une colonie d’abeilles ne produit pas de mâles tant que la colonie n‘a pas suffisamment de ressources et d’ouvrières adultes pour que puisse croître la colonie. Les nouvelles colonies freinent la production précoce de mâles en ne construisant pas de cellules de mâles dans une période de 22 jours en moyenne après la constitution de l’essaim. Ensuite les nouveaux essaims produisent une surface moyenne de 8 % de la surface totale des rayons construits en cou vain de mâles. En comparaison, les colonies qui sortent de l’hivernage produisent une surface moyenne de 13 à 17 % de la surface totale des rayons construits en couvain de mâles. Il y a aussi un facteur de vitesse qui entre en jeu. Les plus gros essaims construisant plus vite une plus grande proportion de cellules de mâles que les petits essaims. Une fois que la colonie a atteint une taille importante (plus de 12 000 ouvrières adultes), le nombre de mâles élevés n’augmente pas proportionnelle ment. Il y a donc a priori une limite maximum à l’élevage de mâles dans les colonies.
La disponibilité alimentaire
Cette limite est logique : il est plus coûteux pour une colonie de produire des mâles que des ouvrières. La disponibilité en nourriture (qualité et quantité) est un facteur de régulation de la quantité de mâles. De
mauvaises conditions d’alimentation conduisent également la colonie à diminuer la production d’ouvrières.
Le couvain de mâles
La colonie ajuste la production de couvain de mâles (œufs, larves et pupes) en fonction du nombre de mâles adultes déjà présents dans la colonie par un processus de rétroaction négative. La présence de couvain de mâles serait un facteur inhibiteur de production de mâles supplémentaires. Des études mériteraient d’être approfondies pour mieux comprendre le processus.
La colonie orpheline
Dans une colonie où la reine est présente, elle produit 99,9 % des mâles adultes de la colonie. Dans une colonie où la reine est absente, les ouvrières tenteront d’élever une reine de remplacement. En cas d’échec, la colonie sera orpheline et l’on parlera de colonie bourdon neuse. Les ouvrières développent alors des ovaires de circonstance et pondent des œufs non fécondés. La colonie investit ainsi dans l’élevage de mâles, seule manière qui lui reste de transmettre des gènes. En dépit de l‘oophagie qui se produit dans ces circonstances, la ponte des ouvrières peut entraîner la naissance de 6 000 mâles adultes supplémentaires. La colonie est toutefois condamnée à mourir. Ceci doit donc être plutôt considéré comme un dérèglement.
Le microclimat des colonies
Les faux bourdons semblent très sensibles aux variations et aux excès de température au stade nymphal. Cela a des conséquences sur leur capacité de reproduction. (Voir article « L‘autre moitié du ciel. Des sper matozoïdes ailés... sous-estimés... incompris... » par Giacomo Acerbi p.31). La concentration en dioxyde de carbone joue probablement également un rôle dans leur développement.
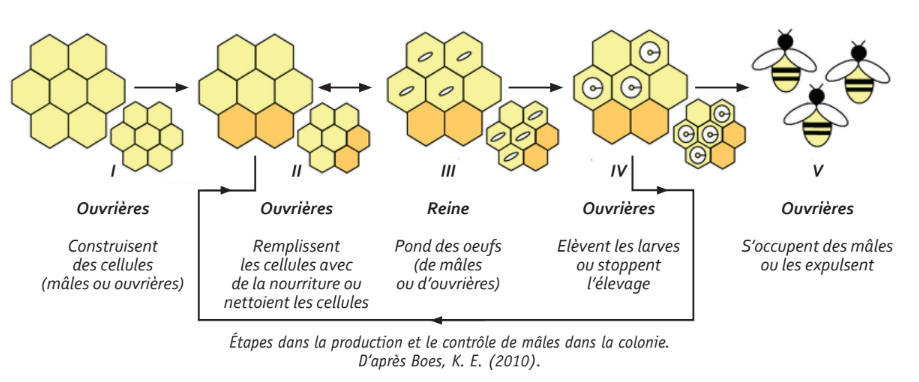
Des étapes pour réguler l’élevage des mâles
À chacune de ces étapes, les abeilles ont la possibilité de modifier l’investissement de la colonie dans l’élevage de mâles en fonction des conditions fixées par l’environnement de la colo nie et sa situation (cellules colorées).
I - Les ouvrières construisent des cel lules. Les cellules de mâles sont plus larges que les cellules d’ouvrières.
II - Une partie des cellules d’ouvrières et de mâles peuvent être utilisées pour stocker nectar et pollen (et de l’eau temporairement) ou nettoyées et réutilisées pour l’élevage de couvain.
III - La reine mesure la taille de la cellule avec ses pâtes antérieures pour pondre des œufs de mâles (non fécondés) dans les plus grandes cellules.
IV - Les ouvrières s’occupent des larves mais ne les conduisent pas for cément toutes jusqu’à l’âge adulte.
V - Les mâles restent dans la colonie jusqu’à leur accouplement avec une reine vierge ou leur expulsion par les ouvrières.
Bibliographie
Boes, K. E.(2010). Honeybee colony drone production and maintenance in accordance with environmental factors : an interplay of queen and worker decisions. Insectes sociaux, 57(1), 1-9.
Bühler, A., Lanzrein, B., & Wille, H. (1983). Influence of temperature and carbon dioxide concentration on juvenile hormone titre and dependent parameters of adult worker honey bees (Apis mellifera L.). Journal of insect physio logy, 29(12), 885-893.
Czekon’ska, K., Chuda-Mickiewicz, B., & Samborski, J. (2015). Quality of honeybee drones reared in colonies with limited and unlimited access to pollen. Apidologie, 46(1), 1-9.
Free, J. B., & Williams, I. H.(1975). Factors determining the rearing and rejection of drones by the honeybee colony. Animal Behaviour, 23, 650-675.
Hwang, J., Lee, J. S., & Kim, H. (2019). Perceived innovativeness of drone food delivery services and its impacts on attitude and behavioral intentions : The moderating role of gender and age. International Journal of Hospitality Management, 81, 94-103.
Lee P.C. and Winston M.L. 1985. The effects of swarm size and date of issue on comb construction in newly foun ded colonies of honey bees (Apis mellifera L.). Can. J. Zool. 63 : 524–527
Li, Z., Huang, Z. Y., Sharma, D. B., Xue, Y., Wang, Z., & Ren, B. (2016). Drone and worker brood microclimates are regulated differentially in honey bees, Apis mellifera. PloS one, 11(2), e0148740.
Page R.E. and Metcalf R.A. 1984. A population investment sex ratio for the honeybee (Apis mellifera L). Am. Nat. 124 : 680–70
Page, R. E., & Erickson, E. H.(1988). Reproduction by worker honey bees (Apis mellifera L.). Behavioral ecology and sociobiology, 23(2), 117-126.
Ratnieks, F. L., & Visscher, P. K.(1989). Worker policing in the honeybee. Nature, 342(6251), 796-797.
Rowland, C. M., & McLellan, A. R.(1987). Seasonal changes of drone numbers in a colony of the honeybee, Apis mellifera. Ecological modelling, 37(3-4), 155-166.
Seeley, T. D., & Mikheyev, A. S.(2003). Reproductive decisions by honey bee colonies : tuning investment in male production in relation to success in energy acquisition. Insectes sociaux, 50(2), 134-138.