Les auteurs, Mallinger, Gaines-Day, et Gratton, examinent les publications relatives aux trois types d’effets que pourraient avoir les espèces gérées sur les espèces sauvages : la compétition alimentaire (1), les changements provoqués dans les communautés végétales (2), la transmission d’agents pathogènes (3). Nous aborderons les points 2 et 3 dans les prochains numéros d’Abeilles & Cie. Examinons ici les études des effets potentiels de compétitions des abeilles gérées sur les abeilles sauvages dans le contexte des écosystèmes naturels (sans tenir compte des écosystèmes gérés, semi-naturels, agricoles ou des terrains expérimentaux).

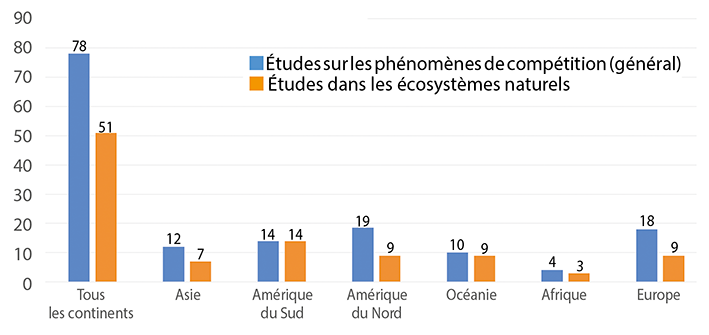
Premier constat, sur les 78 études recensées à l’échelle mondiale entre 1900 et 2016, 51 publications sont relatives à l’étude des phénomènes de compétition alimentaire dans les écosystèmes naturels.
Parmi ces études, neuf concernent des écosystèmes naturels en Europe et l’une d’entre elles étudie l’impact de Bombus terrestris dalmatinus sur Bombus terrestris audax. Il reste donc huit études qui évaluent l’impact d’Apis mellifera sur les espèces sauvages.
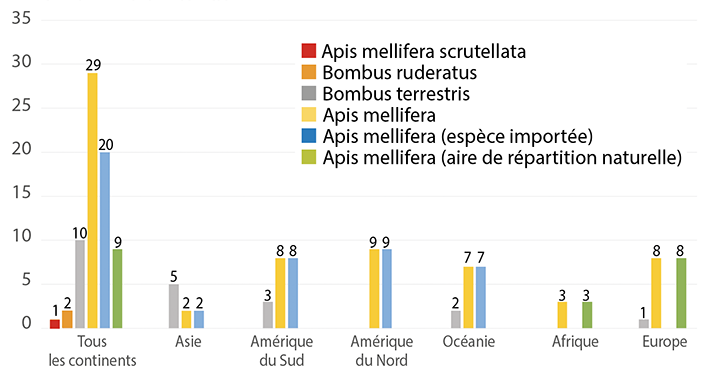
Il n’est peut-être pas inutile de rappeler que Apis mellifera est dans son aire de répartition naturelle en Europe (comme en Afrique).
Aucune de ces huit études ne conclut à une simple compétition positive avec les espèces sauvages. Trois études concluent à une compétition négative : une en Espagne (Torné-Noguera et al. 2016), une en Autriche (Neumayer, 2006), et une en Allemagne (Hudewenz & Klein, 2013). Torné-Noguera et al. ont mesuré la consommation des ressources florales en romarin et thym dans la garrigue du Parc Naturel du Garraf. L’étude conclut que les abeilles mellifères sont les principaux insectes consommateurs de pollen et nectar des deux plantes et que la densité de 3,5 ruches/km2 du parc affecte la communauté des abeilles sauvages. Neumayer a placé deux ruches à 1.650 m d’altitude pendant une saison dans une vallée des Alpes autrichiennes dépourvue d’abeilles mellifères. L’étude conclut que la présence d’abeilles domestiques entraîne une diminution significative des autres pollinisateurs dans le voisinage des ruches tout au long de la saison. Hudewenz & Klein ont étudié les pollinisateurs sur la bruyère callune dans la lande de Lüneburger en Allemagne. Ils constatent une diminution des visites d’abeilles sauvages sur les fleurs en présence des abeilles mellifères. Ils n’ont plus observé aucun effet de cet ordre lorsque les ruches sont distantes de 1.229 mètres.
Trois études arrivent à une conclusion neutre : deux en Allemagne (Kühn et al. 2006 ; Steffan-Dewenter, & Tscharntke, 2000) et une en Grèce (Goras et al., 2006). Kühn et al. ont étudié le cas de Megachile lapponica, abeille oligolectique sur épilobes. 15 ruches d’abeilles mellifères ont été déposées sur le site d’étude sans qu’il y ait une diminution de la fréquence des visites des épilobes par Megachile lapponica, pas plus qu’une augmentation de la durée des voyages à la recherche de nourriture. Aucune diminution du nombre de cellules incubées construites et approvisionnées par Megachile lapponica n’a pu être enregistrée non plus. Steffan-Dewenter, & Tscharntke ont étudié le rayon d’alimentation des abeilles mellifères, la compétition alimentaire et les possibles effets sur la diversité et l’abondance des espèces d’abeilles sauvages dans 15 prairies calcaires d’Europe centrale. Un chevauchement des ressources de 45,5 % a été relevé. Ni le nombre d’espèces d’abeilles ni le nombre de cellules de couvain n’ont été corrélés de manière significative avec la densité des abeilles mellifères.
Goras et al. ont étudié les effets possibles des abeilles mellifères sur le comportement alimentaire des abeilles sauvages sur les fleurs de Cistus creticus dans le nord de la Grèce. Ils ne relèvent aucun effet significatif même avec des densités d’abeilles supérieures à la moyenne européenne de 3,1 colonies/km2.
Deux études ont des résultats mixtes, neutre et négatif en Grande-Bretagne (Forup et Memmot, 2005) et neutre, négatif et positif à l’échelle de l’Europe (Nielsen et al., 2012).
Forup et Memmot s’intéressent à l’impact d’Apis mellifera sur les espèces de bourdons dans les landes du Sud de l’Angleterre. Ils concluent que cet impact est un phénomène complexe. Si la compétition entre les deux espèces n’est pas exclue, leur déclin est, selon eux, imputable à d’autres facteurs. Nielsen et al. relèvent quant à eux une relation positive entre les fréquences de visite des abeilles mellifères et des bourdons, tandis que les relations avec les syrphes et les abeilles solitaires varient (positive, négative et neutre) en fonction de l’espèce végétale étudiée.
On peut facilement conclure que les études que nous venons de présenter, les seules ayant été publiées concernant le continent européen, portent souvent sur des contextes environnementaux ou des espèces animales et végétales spécifiques. La complexité du dossier permet difficilement d’être catégorique dans un sens comme dans un autre concernant les phénomènes de compétition alimentaire dans les écosystèmes naturels. Des études supplémentaires sont nécessaires, en particulier pour le territoire de l’Europe occidentale. Pour compléter le dossier de la coexistence des espèces pollinisatrices, nous envisagerons également l’état des connaissances concernant l’effet potentiel des abeilles gérées sur les abeilles sauvages via des changements dans les communautés végétales et via la transmission potentielle d’agents pathogènes.
Références
- Mallinger, R. E., Gaines-Day, H. R., & Gratton, C. (2017). Do managed bees have negative effects on wild bees ? : A systematic review of the literature. PloS one, 12(12), e0189268.
- Ings, T. C., Ward, N. L., & Chittka, L. (2006). Can commercially imported bumble bees out‐compete their native conspecifics ?. Journal of Applied Ecology, 43(5), 940-948.
- Torné-Noguera, A., Rodrigo, A., Osorio, S., & Bosch, J. (2016). Collateral effects of beekeeping : impacts on pollen-nectar resources and wild bee communities. Basic and applied ecology, 17(3), 199-209.
- Neumayer, J. (2006). Einfluss von Honigbienen auf das Nektarangebot und auf autochthone Blütenbesucher. Entomologica Austriaca, 13, 7-14.
- Hudewenz, A., & Klein, A. M. (2013). Competition between honey bees and wild bees and the role of nesting resources in a nature reserve. Journal of Insect Conservation, 17(6), 1275-1283.
- Kühn, J., Hamm, A., Schindler, M., & Wittmann, D. (2006). Ressourcenaufteilung zwischen der oligolektischen Blattschneiderbiene Megachile lapponica L.(Hymenoptera, Apiformes) und anderen Blütenbesuchern am schmalblättrigen Weidenröschen (Epilobium angustifolium, Onagraceae). Mitteilungen der Deutschen Gesellschaft für Allgemeine und Angewandte Entomologie, 15, 389-392.
- Steffan-Dewenter, I., & Tscharntke, T. (2000). Resource overlap and possible competition between honey bees and wild bees in central Europe. Oecologia, 122(2), 288-296.
- Goras, G., Tananaki, C., Dimou, M., Tscheulin, T., Petanidou, T., & Thrasyvoulou, A. (2016). Impact of honeybee (Apis mellifera L.) density on wild bee foraging behaviour. Journal of Apicultural Science, 60(1), 49-62.
- Forup, M. L., & Memmott, J. (2005). The relationship between the abundances of bumblebees and honeybees in a native habitat. Ecological Entomology, 30(1), 47-57.
- Nielsen, A., Dauber, J., Kunin, W. E., Lamborn, E., Jauker, B., Moora, M., ... & Settele, J. (2012). Pollinator community responses to the spatial population structure of wild plants : A pan-European approach. Basic and Applied Ecology, 13(6), 489-499.