Des petites tranches de ciel... la zone de rassemblement des mâles
Giacomo Acerbi - Traduction : Etienne BRUNEAU
Poursuivons notre « voyage bourdonnant » sur l’évolution des connaissances de certaines caractéristiques comportementales importantes du faux-bourdon, avec ses activités préparatoires et relatives à l’accouplement, la façon dont se manifestent les aires de rassemblement et enfin, leur rôle dans la diffusion et la spécialisation des abeilles et autres hyménoptères sur terre.
C’est un gangbang ? Non !
La zone de rassemblement des mâles (DCA = Drone Congregation Area) est tout sauf un banal « rassemblement de mâles ». C’est en fait le lieu spécifique où des milliers de faux-bourdons et de reines vierges se rencontrent et où la fécondation a lieu. Il s’agit donc principalement d’un comportement social, une dynamique typique et presque exclusive d’Apis mellifera et de certaines espèces de mélipones polyandres : Scaptotrigona (Paxton 2000), Trigona Collina (Cameron 2004)... Il se déroule dans certaines conditions environnementales : dans un espace de 30 à 200 mètres de diamètre (Ruttner 1965), généralement à 15 à 40 mètres au-dessus du sol (Ruttner 1966).
Différentes générations de bourdons de la ruche fréquentent la même DCA chaque saison. Mais les mâles d’une colonie peuvent préférer différentes zones de rassemblement, près de la ruche. Une même DCA peut être visitée par des bourdons de différentes sous-espèces (Ruttner 1972), mais à des moments différents (Benstead 2009).
La DCA se place dans des limites aériennes très précises : les faux-bourdons ignorent totalement la phéromone des reines situées à quelques mètres en dehors de cette zone circonscrite (Ruttner et Ruttner 1965). De plus, les colonies présentes dans un même rayon restent fidèles à la même DCA : différentes générations de bourdons d’une même ruche fréquentent la même DCA chaque saison (Laidlaw et Page 1984). En même temps, les mâles d’une colonie peuvent se répartir dans différentes DCA entourant leur colonie : on a enregistré leur présence jusqu’à dans 10 zones distinctes de rassemblement (Ruttner 1975). Chaque ruche tente de participer à l’avenir génétique de différentes populations d’abeilles, selon une stratégie de survie précise : éviter l’endogamie*1 et maximiser la variabilité génétique. De plus, la présence d’une DCA au même endroit pendant plus de 50 ans a été enregistrée (Jean Prost 1957).
Un lieu de rassemblement peut être considéré comme une DCA s’il est fréquenté par au moins un millier de mâles et si des faux-bourdons sont vus formant des comètes ; et enfin si sa présence a été enregistrée pendant au moins deux jours non consécutifs, séparés par au moins deux semaines (Loper 1992). Le nombre de mâles dans une DCA peut varier en fonction des conditions climatiques (température et vent) et de la densité des colonies dans un territoire d’environ 5 km de rayon (Ruttner 1976). Le nombre de faux-bourdons dans une DCA, dans les zones à forte densité de colonies, varie de 2.145 à 11.750 faux-bourdons (présence simultanée moyenne sur une heure) avec un maximum enregistré de 15.290 (Koeniger 2005).
L’orientation pour l’identification de DCA est liée non seulement à la reconnaissance des caractéristiques paysagères et morphologiques du territoire, mais aussi à une sensibilité spécifique des abeilles au géomagnétisme terrestre.
L’apparition ou non d’une zone de rassemblement pendant la saison de reproduction peut être conditionnée par une multitude de facteurs : conformation du territoire, abri des vents, distance des ruches d’origine des mâles, rayonnement solaire et, enfin, par les caractéristiques comportementales et éthologiques spécifiques des bourdons qui y participent : leur capacité à s’orienter, à voler, à communiquer... Outre ces aspects attitudinaux qui peuvent influencer le choix ou non d’une zone donnée comme DCA, interviennent également des éléments discriminants de nature purement géographique et morphologique d’un territoire. Ces derniers sont liés à sa conformation, à la présence ou non de cours d’eau, de bois, de zones urbanisées, de champs.
Les recherches et les expériences de terrain ont montré que les DCA se trouvent à proximité de terrains ouverts entourés d’arbres ou de végétation très haute et que la présence de cours d’eau, les intersections de routes, la présence de grands « obstacles » (par exemple un arbre centenaire...), l’alternance de zones sans végétation... sont probablement parmi les facteurs qui facilitent grandement l’orientation des mâles et des reines vierges et contribuent donc à l’existence des DCA (Loper 1992). De manière surprenante, dans les résultats de la recherche menée par Galindo-Cardona, dans 71 % des DCA il y avait une portion de territoire avec une couverture urbaine : cela peut suggérer que les mâles et les reines vierges utilisent les maisons, les bâtiments, les routes... comme points de référence pour s’orienter.
En outre, la majorité des zones examinées dans lesquelles une DCA a été détectée présentent un abri contre les vents soufflant du nord et sont caractérisées par une ligne d’horizon libre et ouverte, avec peu de points de référence (10 %) et normalement avec un terrain sous-jacent plat, à pente maximale de 19 %.
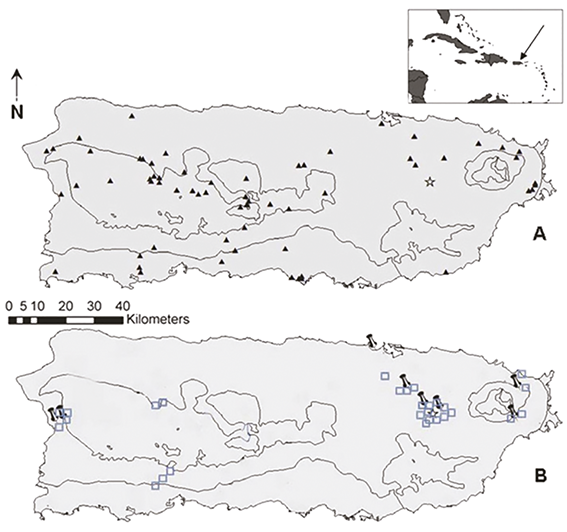
La figure A montre la carte de Porto Rico avec les ruchers identifiés présents sur l’île (triangles noirs) et notre rucher (étoile). La figure B montre où les DCA sont présents (n° 8 et identifiés par un point noir) et où ils sont absents (carrés bleus) - d’après l’étude de Galindo-Cardona 2012
L’important et fascinant sens de l’orientation des abeilles résulte non seulement de la reconnaissance des caractéristiques paysagères et morphologiques du territoire, ainsi que de la distribution de la lumière et de la direction du vent (Hempel 2009), mais aussi d’une sensibilité spécifique au géomagnétisme terrestre (Hsu 2007). C’est le fascinant phénomène de magnétoréception*2 (Yoshi 2009) grâce auquel tant d’espèces vivantes se situent dans l’espace, s’orientent, « tracent des routes » pour voyager, revenir, s’accoupler, se reproduire, subvenir aux besoins de leur progéniture, etc. De nombreuses études ont confirmé que les abeilles, ainsi que les faux-bourdons, disposent d’une boussole interne pour identifier et tracer des itinéraires dans la bonne direction (Merlin 2011), en se basant également sur la distribution de la lumière et la direction du vent (Hempel 2009). L’aptitude des mâles, basée sur le rythme circadien*3 exprime et englobe de nombreuses capacités de « navigation ». Une étude (Galindo-Cardona 2012), qui porte sur les caractéristiques paysagères des DCA, met en évidence de nombreux aspects de la conformation territoriale d’une zone de rassemblement, mais révèle surtout que, dans le choix de la localisation géographique d’une DCA, l’éthologie des mâles joue un rôle crucial : leur capacité à voler, à s’orienter... et leur prédilection pour aller « au sud » ! En effet, les localités dans lesquelles les zones de rassemblement ont été trouvées ont une conformation territoriale avec une orientation sud. Les faux-bourdons utilisent le soleil comme boussole (von Frisch 1967) et volent dans une direction sud/sud-est en fonction du rayonnement solaire (McCune et Keon 2002).
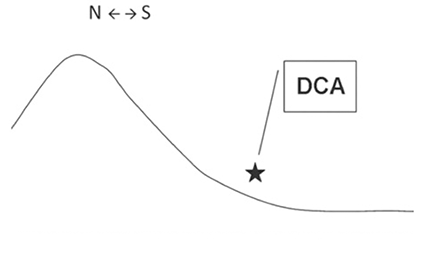
Diagramme d’une section topographique représentant la pente et la conformation orientée vers le sud d’une DCA. L’étoile indique l’emplacement exact de la DCA. N et S représentent le Nord et le Sud - d’après l’étude Galindo-Cardona 2012
Reprise de l’étude Galindo-Cardona
Qui vient en premier ? Les reines vierges ou les mâles ?
Les bourdons s’assemblent et forment des DCA indépendamment de la présence ou de l’absence de reines vierges (Ruttner 1966, Jean Prost 1957), la présence de femelles n’est pas cruciale pour leur formation, même dans ce cas ... le plus beau sexe est ... attendu (Koeniger 2004).
Les points de référence importants pour le rassemblement des mâles (Ruttner 1985) sont la ligne d’horizon (montagnes, cime des arbres...), les anomalies du champ magnétique terrestre, etc. Mais la production de phéromones par les mâles pour attirer d’autres mâles est fondamentale. Des études récentes ont permis d’établir qu’un bouquet de substances volatiles, probablement émises par les glandes labiales ou celles situées dans les antennes, constitue un signal « odorant », peut-être fondamental pour la constitution des zones de rassemblement (Bastin 2017). L’émission d’une gamme de signaux olfactifs par les bourdons pourrait expliquer à la fois les frontières géographiques très limitées des DCA, et aussi la façon dont les reines vierges reconnaissent et sont attirées par les zones encombrées de bourdons (Bastin 2017).
La compétition. A vos marques, prêts... partez !
Les bourdons créent des formations de vol dans la DCA, avec une forme similaire à celle des comètes, ainsi appelées » comètes à faux-bourdons « ou « comètes de fécondation » (Koeniger 2005).
Elles peuvent apparaître et se dissoudre en quelques secondes (Gary 1962) et déterminent le comportement reproductif le plus important des abeilles : c’est ici que se décide la « pole position » et, comme dans une course automobile, les éléments fondamentaux sont : l’accélération, la vitesse, la capacité à maintenir une position dominante sur les autres ... La course, la compétition pour l’accouplement ne permet pas de marge d’erreur, c’est ici qu’elle est déterminée et qu’elle discrimine qui transmettra ou non ses gènes à la descendance. Le nombre total de mâles « en compétition » au sein d’une comète varie de 20 à 41 (Koeniger 2005), avec un poids unitaire (de chaque spécimen individuel) allant de 79 mg à 223 mg, avec des vitesses de vol moyennes allant de 2,6 mètres/seconde à 4,6 mètres/seconde, et une capacité d’accélération de 10 mètres/seconde (Koeniger 2005).
Le drone capable d’exprimer une plus grande forme physique et une plus grande vigueur est reproduit par la compétition aérienne qui combine force, stratégie, vitesse et vigilance.
René Magritte, Golconde
La durée moyenne du séjour d’un faux-bourdon dans la comète varie entre 0,7 et 1,7 secondes : ils entrent et sortent continuellement de la formation de vol. En fait, l’espace de temps pour atteindre une position « intéressante » pour l’accouplement est très limité et le roulement continu dans la comète peut peut-être être interprété comme la perte momentanée de l’espoir de se placer dans une position gagnante et la tentative de le regagner. Cela a d’importantes implications de nature génétique et héréditaire : le sujet capable d’exprimer une plus grande forme et une plus grande vigueur physique se reproduit et, chez les mâles, c’est déterminé par une compétition aérienne qui combine force, stratégie, vitesse et disponibilité. Cela confirme que l’aptitude au vol des bourdons et toutes les caractéristiques qui y sont liées est l’un des éléments clés déterminant l’hérédité dans la descendance et la capacité de reproduction des abeilles. La distance entre la reine vierge et les bourdons dans la comète varie de 4 à 15 cm, mais seuls ceux qui parviennent à s’approcher à moins de 10 cm, en volant à sa hauteur et en restant dans une orbite de 2.000 centimètres cubes, pourront peut-être l’attraper (Koeniger 2005). Aucun contact physique entre les bourdons n’a été observé, mais ils ajustent plutôt leur position à celle de leurs voisins tout en la défendant, et dans 73 % des cas, ils commencent à poursuivre la reine en groupe : les combats et les collisions en vol peuvent entraîner la perte de la trajectoire de la reine (Koeniger 2005). Cependant, le comportement des bourdons dans les comètes est également déterminé par les différentes étapes de l’accouplement.
Pendant la parade nuptiale, la reine vole rapidement et atteint une certaine hauteur au-dessus du sol (Koeniger 1989) : à partir de là, la « poursuite » commence et culmine avec l’un des faux-bourdons « bien placés » qui attrape la reine vierge et s’accouple ; pendant cette phase, il y a une réduction considérable de la vitesse qui permet aux autres faux-bourdons d’essayer d’obtenir une meilleure position dans la comète.
La fécondation a lieu dans l’air à une hauteur de 15 à 60 mètres (Loper 1992), à une vitesse qui peut atteindre 12 Km/h (Oertel 1956).
La durée de la copulation est inférieure à deux secondes et, probablement pas par hasard, cet intervalle de temps correspond à la fréquence moyenne de séjour d’un mâle à l’intérieur d’une comète (0,7 -1,7 seconde).
Dans la frénésie, des erreurs peuvent se produire, et la poursuite dans une position de pré-copulation et la prise entre les bourdons eux-mêmes est un événement fréquent (Gary 1963). Cela peut être causé par une fausse perception visuelle (Gary 1963) due, par exemple, à la distance de la reine vierge dans la comète.
Sans les mâles ? Que deviendraient les abeilles ?
Ces petites « portions de ciel » garantissent la reproduction et la survie des abeilles depuis des millions d’années, grâce à leur rôle précieux dans le développement et le maintien de la variabilité génétique dont les DCA sont le berceau et dans laquelle les faux-bourdons jouent un rôle crucial. Une reine peut être fécondée par 6 à 28 mâles avec une moyenne statistique d’environ 15 (Holm 2010) ; un levier simple et en même temps excellent pour contrebalancer la parthénogenèse*4 et assurer une grande hétérogénéité de la descendance. La complexité, la consommation considérable d’énergie, l’investissement de fonctions biologiques spécifiques et particulières, la capacité de se conformer aux tendances environnementales... constituent une « dynamique » et un « phénomène » articulés qui sont à la base du succès adaptatif d’Apis mellifera, de son extraordinaire longévité et de sa présence généralisée à différentes latitudes sur cette planète.
Le retour aux anciennes traditions de l’art populaire baroque en Allemagne et en Pologne. Ruches sculptées : la force vitale de la fertilité des abeilles est le thème principal de la représentation figurative.
Identifier, connaître et préserver les colonies d’abeilles peut être très utile pour : étudier les différentes populations d’abeilles d’un territoire donné (Loper 1992), permettre leur caractérisation en fonction de leur diversité génétique (Collet 2009), estimer leur structure génétique*5 (Collet 2009), identifier la présence de maladies ou d’éventuelles immunités adaptatives*6 dans les colonies d’un territoire donné grâce à l’étude des faux-bourdons comme échantillons de la variabilité génétique d’une zone donnée (Evans 2006, Robinson 2008). Connaître les aspects du comportement reproducteur des abeilles, identifier comment et où il se produit, est fondamental pour avant tout, protéger ces précieuses zones de la pression anthropogénique (produits agrochimiques, perte de biodiversité etc...).
Enfin, la prise en compte et la sensibilisation à l’éthologie des bourdons peut être stratégique et fonctionnelle pour tenter de mettre en œuvre des pratiques apicoles plus efficaces, ponctuelles et fonctionnelles pour la conservation, l’amélioration génétique et la sélection généalogique (Ruttner 1976) : des activités pour lesquelles le rôle génétique et qualitatif de la composante paternelle est la base pour obtenir et consolider des résultats satisfaisants. D’autre part, comme nous l’avons vu, c’est précisément le comportement particulier des bourdons qui exprime l’ensemble des mécanismes inclusifs ou discriminatoires qui permettent... ou non à une ruche de transmettre ses gènes aux générations futures. C’est dans la capacité de vol, d’orientation, de vitesse…, dans les qualités physiques et reproductives de ce « sperme ailé » que déterminent la composition, l’empreinte et l’impact génétique d’une population d’abeilles. Qui sont donc les faux-bourdons ? Ce sont comme les reines, des « photocopieurs », mais dont toutes les copies sont différentes les unes des autres, comme les spermatozoïdes chez les autres êtres vivants.
« Le royaume des abeilles »
Sa Majesté a décidé de se multiplier sereinement (traduit du vieil allemand) Circa. 1851, Carl Vogt, dans Études des États Animaux.
Vocabulaire : Drone
Un mot en vogue aujourd’hui. Ce n’est que depuis 1946 que le mot « drone » désigne aussi, en anglais, les avions radiocommandés sans pilote. Jusqu’alors, il signifiait simplement « bourdon », c’est-à-dire mâle d’abeille, et tout au plus, comme les bourdons ne font pas de miel, il signifiait « paresseux, paresseuse ». Cependant, on ne sait pas pourquoi le nom de bourdon a été choisi ; probablement en raison du malentendu selon lequel les bourdons étaient l’élément le plus sacrifiable de la colonie comparé à la femelle. Un objet volant sans pilote à bord, avec la possibilité de transporter des charges, a été exploité pour la première fois en guerre en 1849, pour bombarder Venise. Un des officiers du général autrichien Von Radetzky, a eu l’idée de lancer une attaque avec des ballons aérostatiques lancés depuis un navire à l’ancre, sans équipage et avec une quinzaine de kilogrammes d’explosifs. Un dispositif de chronométrage rudimentaire utilisant du charbon de bois et du fil d’amorçage en coton aurait libéré les bombes au-dessus de Venise, mais... des conditions météorologiques défavorables et des vents irréguliers ont fait que la plupart des ballons sont retournés vers les lignes autrichiennes.
Dans le jargon musical, le bourdon, quant à lui, indique une note d’accompagnement ou un accord continu, suscitant divers effets. Les plus célèbres sont ceux du Scherzo de la Pastorale de Beethoven et de la Finale de la Symphonie 104 de Haydn. En Europe continentale, ce type de son est connu sous le nom de « bourdon » : son origine rappelant ou imitant un son grave et continu.
Les premiers « drones », utilisés en 1849 par l’armée de Radetzky pour tenter de bombarder Venise
1. Endogamie : la reproduction sexuée entre individus du même noyau parental, donc consanguine.
2. Magnétoréception : une sorte de boussole biologique présente chez certaines espèces vivantes : de nombreux oiseaux, homards, baleines, dauphins, requins, raies manta, abeilles, ainsi que des micro-organismes et certaines plantes. Il permet à ces formes de vie de détecter le champ magnétique terrestre, de créer des cartes territoriales, avec l’altitude relative, la position, la direction et donc de s’orienter.
3. Rythme circadien : l’horloge biologique complexe qui détermine, chez les animaux et les humains, une série de stimuli synchronisants innés (sommeil-veille/production d’hormones...) dans les 24 heures d’alternance jour/nuit.
4. Parthénogenèse : forme de reproduction sexuelle des plantes ou des animaux qui ne nécessite pas de fécondation, amphigonique ; elle ne nécessite pas l’union de gamètes mâles et femelles.
5. Structure génétique : la constitution génétique d’une population en termes quantitatifs (fréquences alléliques et phénotypiques) et qualitatifs (variants alléliques dans une population).
6. Immunité adaptative : capacité acquise du système immunitaire à réagir, basée sur la formation de cellules mémoire dans l’organisme pour une réponse rapide à un antigène.