Il y a quelques années, certains chercheurs ont noté qu‘il y avait une population d‘abeilles varroa-tolérantes en Norvège dans les ruchers de Terje Reinertsen.
Leur caractéristique est que vous n‘avez pas du tout besoin de les traiter contre les varroas, même s‘il y en a des quantités variables au cours d‘une saison. Et cela fait plus de 20 ans. Aujourd’hui, en Belgique et en France, on parle énormément du caractère appelée VSH (Varroa Sensitive Hygiene). En d’autres mots, des abeilles qui ont la capacité de nettoyer les cellules de couvain dans lesquelles les acariens se reproduisent. VSH n‘est cependant pas une caractéristique importante des abeilles norvégiennes tolérantes. On ne peut cependant pas nier que le comportement de réoperculation n’est pas lié au VSH. Lors de son travail de doctorat, Mélissa Oddie une chercheuse canadienne a découvert un trait important présent chez les abeilles de Terje, le comportement de désoperculation et de réoperculation des cellules dans lesquelles les femelles varroas se reproduisent. L’ouverture de l’opercule perturberait apparemment la reproduction de l‘acarien.
TERJE REINERTSEN, PÈRE DE L’ABEILLE TOLÉRENTA NORVÉGIENNE
Texte réalisé sur base d’un article d’Erik Ijsterlund dans la revue Gadden 1/2020 et de notre rencontre lors du congrès des apiculteurs suédois à Sunne (Biodlarkonferensen 2020

C’est en 1993, que les premiers varroas ont été identifiés dans certains ruchers en Norvège. Comme partout, décision avait été prise qu‘il était essentiel de les combattre. Les autorités se sont alors basées sur ce qui était préconisé dans les pays qui avaient une plus longue expérience avec l‘acarien. Entre autres choses, il a été recommandé d’utiliser dans les ruches des plaques imbibées d‘acide formique. Terje a suivi ces conseils et a traité ses abeilles de cette façon en 1994. Il a perdu 70 % de ses colonies au cours de l‘hiver 1994-95 et d‘autres ont perdu 100 % de leurs abeilles. Comme traitement de contrôle au printemps, une seconde application d‘acide formique a été recommandée mais avec moins de trous dans le plastique qui recouvre la plaque imbibée d‘acide formique afin que l‘évaporation ne soit pas aussi forte. Terje l‘a utilisé sur les colonies survivantes. Après avoir analysé tous les résultats de cette lutte, Terje et quelques autres ont conclu qu’en perdant autant d‘abeilles en combattant l‘acarien, on pouvait tout aussi bien ne pas traiter, sans en perdre davantage. La question était alors « comment favoriser une meilleure tolérance des abeilles au varroa de façon plus naturelle et moins destructrice ? ».
Bases du changement
Terje suspectait un impact important des traitements sur la résistance des abeilles et sur leur comportement. A l’époque, on ne savait que peu de choses sur l‘importance du système immunitaire face aux parasites et maladies et de l’importance de maintenir un bon équilibre entre les différents micro-organismes (bactéries, virus et autres - microbiome) présents dans les êtres vivants. De nombreux chercheurs travaillent aujourd’hui sur le microbiome, y compris chez les abeilles. On sait que les produits chimiques déséquilibrent ce microbiome générant des dysfoncionnements. La défense contre les microbes nuisibles, par exemple contre les virus, en est affaiblie.

Lors de son travail de sélection, il a pu constater que certains comportement étaient liés à la génétique mais qu’on sous-estimait probablement le comportement d’apprentissage qu’il a pu observer à de nombreuses reprises.

Achat de colonies jamais traitées
En 1995, Terje a remplacé ses pertes en achetant plus de 30 colonies.
Celles-ci provenaient d‘une zone où le varroa n‘était pas encore présent, de sorte que ces colonies d‘abeilles n‘avaient jamais été traitées contre l‘acarien. Certaines de ces colonies d‘abeilles n‘étaient que des ruchettes de division qui avaient élevé de nouvelles reines au départ de leur propre couvain. Les colonies achetées sont finalement devenues de bonnes ruches. Elles étaient plus nombreuses que celles qui avaient survécu au varroa et à l‘acide formique. Celles-ci n‘étaient pas aussi fortes mais étaient intéressantes. À partir de l‘hiver 1995-1996, les pertes sont redevenues acceptables, malgré l‘absence de traitement.
En raison des pertes importantes d‘abeilles chez beaucoup d’apiculteurs, la recontamination d‘acariens n‘était plus assez forte pour affaiblir les abeilles non traitées. C’est à cette époque que Terje a commencé à élever des reines, ce qu’il fait toujours aujourd’hui. Il a cherché différentes manières d‘identifier les colonies d‘abeilles qui ne developpaient pas de populations d‘acariens importantes et qui présentaient une bonne résistance aux virus. Après quelques années d’expérimentation où les colonies trop chargées en varroas étaient systématiquement suprimées, il a constaté des changements significatifs. Durant les toutes premières années, il perdait environ 30 % de ses colonies et assez rapidement, ce chiffre s’est réduit pour ne plus atteindre aujourd’hui que 3 à 5 % de pertes hivernales.
Programme d’élevage
Il remplace les reines des plus mauvaises colonies et il réalise un élevage au départ des meilleures colonies. Il ne remplace pas systématiquement les vieilles reines. Il ne le fait que lorsque les colonies d‘abeilles sont mauvaises, que les reines soient jeunes ou âgées. Les colonies sont heureuses de changer leur reine elles-mêmes. Les jeunes reines vierges sont de préférence fécondées au centre d‘une région où il peut s’assurer d‘avoir de bonnes colonies. Il travaille avec les populations locales, et évite un élevage intensif, la consanguinité ou l’utilisation de stations ne disposant que d’un petit groupe de colonies à mâles. De plus, l‘insémination n‘est pas utilisée. Ces dernières alternatives peuvent parfois être nécessaires pour empêcher l‘épuisement des bons caractères dans une population, en fonction de la proximité d‘autres types d‘abeilles. C’est ainsi qu’une vitalité et une variation génétique élevée sont maintenues dans l’environnement des colonies de Terje. Initialement, il ne pensait pas pouvoir arriver à un tel résultat avec le nombre d‘abeilles qu‘il avait et il a essayé d‘autres lignées de Buckfast et quelques autres variétés d‘abeilles. Aujourd‘hui, il limite le patrimoine génétique de ses abeilles à celui de sa région.
Autres abeilles résistantes
Il y a dix ans, il a acquis 14 reines norvégiennes Carnica accouplées dans une zone d‘accouplement de rennes en Norvège. Ces reines ont été ajoutées à ses colonies.
Les colonies ont été placées chez un apiculteur à environ 60 km de son exploitation. Elles sont également assez isolées des autres abeilles. Aujourd‘hui, cette population constitue une centaine de colonies. Toutes les nouvelles reines sont fécondées à cet endroit. Ces colonies ne sont jamais traitées contre le varroa. Cet apiculteur a toujours subi de très faibles pertes hivernales. Vous pouvez peut-être le faire chez vous si vous souhaitez obtenir des abeilles varroa-résistantes d‘autres variétés que celles de Terje.
Du côté des chercheurs
Un certain nombre de reines de Terje ont été achetées par plusieurs universités européennes. En 2013 certains essais ont été présentés au professeur Peter Neumann de l’Institut de santé des abeilles de l’Université de Berne en Suisse. Celui-ci s’est rendu sur place en 2014 avec une doctorante Sandra Lanz qui a présenté sa thèse « L’abeille européenne Apis mellifera survivant à Varroa destructor et aux virus », basée sur les abeilles de Terje. Depuis 2015 Melissa Oddie, du même institut, lui a rendu visite plusieurs semaines chaque été pour réaliser sa thèse sur la tolérance de ses abeilles. Celle-ci a été publiée durant l’été 2018. Les autorités norvégiennes ont alloué récemment 8,5 millions de couronnes norvégiennes (1,14 millions €) pour la recherche sur les abeilles de Terje tolérantes aux varroas en Norvège, ce qui va permettre à cette chercheuse de poursuivre son travail sur ces abeilles.
La recherche met en évidence que la sélection de Terje a abouti au développement de 5 traits favorables dans ses colonies : les abeilles sont douces et faciles à travailler, les colonies sont plus populeuses que la normale et la production de miel a augmenté. Leur système immunitaire est fort et elles ont développé une tolérance au varroa.
La zone occupée par les abeilles de Terje s’agrandit
Terje veut toujours aider ses collègues et les débutants, c’est ainsi qu’il organise des cours pour débutants depuis de nombreuses années. Une fois par mois, d‘avril à septembre, l‘association locale se réunit dans son jardin. Il vient en moyenne de 30 à 35 personnes.
Il y a maintenant entre 1 500 et 2 000 colonies Terje en Norvège. Il y a environ 100 apiculteurs qui les conduisent sans traitement contre le varroa. La plupart d‘entre eux se trouvent dans la région où habite Terje.
COMMENT EXPLIQUER CETTE TOLÉRANCE DES ABEILLES PÈRE DE L’ABEILLE NORVÉGIENNE ?

Mélissa Oddie a fait partie de l’équipe de Peter Newman et a été amenée à étudier de plus près le phénomène de tolérance au varroa observé dans le rucher de Terje Reinertsen. Lorsqu’elle a débuté son travail, tout laissait croire que le succès reproductif réduit de Varroa destructor était une condition essentielle pour la survie naturelle des colonies infestées d‘Apis Mellifera. En effet, le succès de reproduction des acariens et les niveaux de population d‘acariens étaient significativement inférieurs dans les colonies norvégiennes survivantes par rapport aux colonies témoins locales sensibles aux varroas. La proportion d‘acariens endommagés comme indicateur indirect de l‘efficacité du comportement de toilettage et de l‘élimination du couvain présentant une reproduction des acariens (VSH) par les ouvrières n‘étaient pas significativement différentes entre les colonies survivantes et les sensibles, ce qui indique que ces deux mécanismes étaient peu susceptibles d‘expliquer la survie naturelle de ces colonies norvégiennes. Cela concorde avec les résultats antérieurs pour Gotland (Suède), où les différences de comportement hygiénique et de toilettage n‘étaient également pas apparentes entre les colonies locales survivantes et sensibles aux acariens. Étant donné que seul le couvain provenant de colonies sensibles a été utilisé pour les expériences dans les colonies hôtes survivantes et sensibles, on ne peut lier le phénomène au couvain, qui pourrait par exemple venir de variations de phéromones émises par le couvain.
Un moindre succès reproducteur

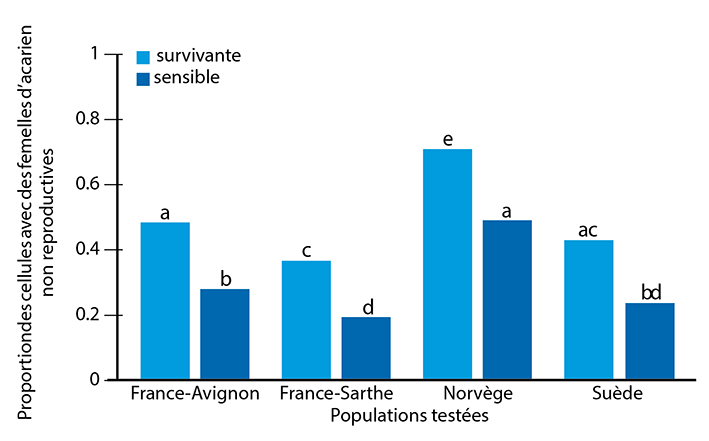
Les niveaux de population d‘acariens, estimés par le nombre de chutes naturelles, étaient significativement plus faibles dans les colonies survivantes que dans les témoins sensibles. Des niveaux d‘infestation inférieurs par les acariens sont une explication évidente de la survie de la colonie et peuvent résulter d‘un succès reproducteur réduit des acariens. En effet, plusieurs exemples le montrent. Seule environ la moitié des acariens dans les colonies de Gotland ont réussi à produire des acariens filles viables qui contribuent à la population d‘acariens de la colonie. Les taux de reproduction étaient d‘environ 0,84 progéniture par fondatrice dans la population norvégienne, c’est à dire un succès de reproduction des acariens réduit d’environ 30 % (graphique 1), suggérant ainsi fortement qu‘une telle réduction est suffisante pour permettre la survie des colonies. Cette réduction du taux de reproduction est également conforme aux études antérieures effectuées sur les abeilles africanisées (0,79) et les abeilles européennes sur une île brésilienne (0,54). De même, le succès de reproduction des acariens dans la population survivante des acariens d‘Avignon a également été réduit de 30 % par rapport aux colonies locales sensibles aux acariens.
L’objectif était donc de comprendre les mécanismes qui conduisent à la baisse du succès de reproduction des acariens dans ces colonies. En d’autres mots, quels sont les traits comportementaux non identifiés des abeilles ouvrières qui semblent suffisants pour expliquer le succès reproductif réduit des acariens et, finalement, la survie des colonies ?
Un travail en quatre étapes
- 1. Inspection des cellules : les ouvrières examinent l’opercule en cire des cellules avec leur langue et leurs antennes.
- 2. Percement des cellules : les ouvrières utilisent leurs mandibules pour ouvrir l’opercule. Ces deux premières étapes sont toujours réalisées dans la même séquence. Une transition majeure se produit après l‘étape 2, où les ouvrières peuvent choisir entre les deux étapes suivantes.
- 3. Réoperculation des cellules : les ouvrières utilisent les sécrétions des glandes cirières et leurs mandibules pour réoperculer les cellules, ce qui fait que le dessous d‘une cellule réoperculée affiche un trou sombre notable avec une cire visuellement mate.
- 4. Enlèvement du couvain : les ouvrières retirent et jettent le couvain des cellules infestées d‘acariens.
- L‘ensemble de la séquence comportementale est flexible en raison de la prise de décision par plusieurs ouvrières impliquées. La séquence peut s‘arrêter après le stade 1. (c‘est-à-dire que les cellules sont étudiées, mais non percées), ou après le stade 2. (c‘est-à-dire que les cellules percées peuvent être réoperculées ou non) et la désoperculation et/ou la réoperculation peuvent n‘être effectuées qu‘en partie.
Désoperculation/réoperculation par les abeilles
Melissa Oddie a mis clairement en évidence lors de ses travaux que la réoperculation est à la fois plus fréquente et ciblée sur les cellules infestées d‘acariens dans toutes les populations étudiées survivant par sélection naturelle par rapport aux témoins sensibles locaux. Cela soutient également l‘idée que la désoperculation/ réoperculation par les abeilles adultes est un mécanisme comportemental atténuant les infestations d‘acariens.

La désoperculation expérimentale des cellules de couvain a montré que la réoperculation peut réduire directement le succès de reproduction des acariens, sans sacrifier les nymphes du couvain. L‘efficacité de la réoperculation comme mécanisme naturel pour réduire le succès de reproduction des acariens a été démontrée en combinant les données des essais de désoperculation expérimentale. l’expérience a démontré que le comportement de réoperculation peut réduire le succès de reproduction des acariens au niveau des colonies et n‘est pas seulement corrélé avec lui. Cette première analyse nécessite cependant une confirmation statistique. Pour l’instant, la seule chose dont on peut être certain, c’est que la réoperculation est directement liée à la réduction du succès de reproduction des acariens. On ne peut pas encore prétendre que ce comportement est le mécanisme moteur de cette effet recherché. Il pourrait être essentiel, comme il pourrait n‘être que le signe d‘un mécanisme différent non encore identifié. Cependant, ce qui est pratiquement certain, c’est que la réoperculation est un bon indicateur comportemental à utiliser en élevage, car il est étroitement lié à la diminution du nombre de descendants d‘acariens et dès lors à la survie naturelle des abeilles européennes infestées par Varroa destructor et non traitée. De même, il n’a pas été possible d’exclure que l‘élimination des cellules infestées et d‘autres facteurs aient également contribué à la survie naturelle des acariens de ces populations. En effet, le phénotype des colonies d‘abeilles mellifères qui survivent naturellement à Varroa destructor est très probablement déterminé par une série de caractères de l‘hôte, les interactions locales génotype-environnement, la variation des pathogènes, la disponibilité des ressources et la gestion apicole. Cependant, en raison de son évolution rapide et parallèle dans les quatre populations survivantes étudiées et de l‘impact significatif sur le succès de reproduction des acariens, la réoperculation semble être un mécanisme clé commun et précédemment ignoré pour la survie des colonies.
Un mécanisme rentable
Le retrait du couvain, bien que potentiellement lié à la réoperculation, ne sera probablement pas un mécanisme principal car la perte d‘individus compromettrait la compétitivité des colonies. De plus, la population norvégienne à l‘étude ne connaît pas de taux accrus de retrait de couvain, ce qui indique qu‘au moins pour cette population, il ne s‘agissait pas d‘un mécanisme de survie important. En revanche, la réoperculation ne nécessite que du temps et de l‘énergie pour manipuler les opercules de cire. Il semble donc que ce soit un mécanisme beaucoup plus rentable que le prélèvement de couvain pour atteindre des objectifs similaires. La reproduction des acariens n‘a pas besoin d‘être complètement interrompue, mais seulement réduite pour affecter suffisamment la croissance de la population pour assurer la survie des colonies. Le moment où les abeilles sensibles aux acariens développent un phénotype de colonie survivant peut actuellement être estimé à un minimum de 17 ans bien que cette période puisse en effet être plus courte. Sur le terrain, dans les populations de Gotland et de Norvège, des différences dans les taux de survie ont été observées en 3 ans environ, et , Terje a réduit de manière significative les taux de perte en 5 ans environ. Malheureusement, aucune de ces données n‘ont fait l’objet de publications scientifiques, mais ce sont les expériences directes des éleveurs. La résistance à la reproduction peut se faire extrêmement rapidement, si l‘on dispose d‘une diversité génétique suffisante. Plus théoriquement, la rapidité de l‘adaptation peut s‘expliquer en quatre points : (1) Le comportement provient très probablement d‘une préadaptation car la désoperculation a été enregistrée dans toutes les populations d‘abeilles mellifères étudiées jusqu‘à présent, (2) la réoperculation et la détection des acariens peuvent s‘expliquer par de simples changements dans les seuils d’activation des ouvrières. Les changements de comportement dans un répertoire d‘immunité sociale peuvent être optimisés progressivement avec un impact considérable au niveau de la colonie. (3) La pénalité liée à la désoperculation d‘une cellule non infestée n‘est pas mortelle pour le couvain, ce qui donne aux erreurs inévitables un impact beaucoup plus faible, et (4) la séquence comportementale détection/désoperculation partielle/analyse/désoperculation ou réoperculation peut être effectuées par différentes ouvrières, ce qui offre de nombreuses possibilités de changements adaptatifs indépendants et non liés. Il est probable que la réoperculation soit un trait commun à de nombreuses populations d‘abeilles, sinon toutes, et que les ancêtres de nos populations survivantes actuelles étaient des colonies individuelles qui présentaient à la fois une fréquence de réoperculation élevée et une sensibilité à la santé du couvain.
À la lumière de ces résultats et de l‘omniprésence de la réoperculation dans les populations d‘abeilles mellifères, cette séquence comportementale semble faire partie intégrante des soins du couvain et de la défense contre les agents pathogènes chez A. mellifera. Il est possible pour les colonies d‘insectes eusociaux de développer des traits d‘immunité sociale à faible coût via de simples changements adaptatifs, qualitatifs et quantitatifs dans le comportement des ouvrières. Dans la perspective d’une apiculture durable, il semble prudent d’évoluer et de pouvoir tirer parti des mécanismes efficaces favorisés par la sélection naturelle pour gérer les maladies infectieuses.

h) - Pourcentage de cellules réoperculées à chaque stade du développement nymphal dans l‘AHB (ligne noire), A. m. scutellata (ligne grise continue) et A. m. capensis (ligne grise en pointillés). Les stades de développement nymphaux suivants sont sl = larves étirées, pw = yeux blancs, po = yeux pâles, pp = yeux roses, pr = yeux pourpres, yt = thorax jaune, gp = coussin gris, et gt / r = thorax gris / repos et le nombre total de cellules ouvertes par stade est également indiqué.
Une confirmation
D’autres auteurs (Martin, S.J., Hawkins, G.P., Brettell, L.E3) travaillant dans des pays plus lointains ont également observé la réoperculation des cellules de couvain dans toutes les colonies, les taux de réoperculation étant les plus bas chez les colonies sensibles au Varroa, et les plus élevés étant systématiquement constatés dans le couvain d’ouvrières de populations résistantes aux acariens du Brésil (AHB) et d‘Afrique du Sud (A . m. scutellata et A. m. capensis). Ils ont également observé que dans le couvain d‘abeilles mellifères africaines envahi par le petit papillon de cire (Achroia grisella), les cellules étaient fréquemment operculées plutôt que les pupes enlevées. De même, 57 % des cellules non operculées dans une colonie fortement infestée par la grande teigne (Galleria mellonella) ont été réoperculées dans les 24 heures suivant la désoperculation. Les colonies de capensis à Pniel étaient uniques pour réoperculer un grand nombre de cellules de mâles. Ce sont toutes des colonies survivantes d‘une épidémie de loque américaine (Paenibacillus larvae). Ils ont observé sur des exemples liées à la petite et à la grande teigne ainsi qu’en cas de loque que le comportement de faire un petit trou dans l’opercule peut être une réponse générale pour permettre une étude plus détaillée de la nymphe en développement (ce qui peut expliquer la faible présence de ce caractère dans les populations sensibles aux Varroas). Après l‘arrivée des acariens, ce comportement semble avoir été coopté et sélectionné dans le cadre d‘un mécanisme de défense contre Varroa ; par conséquent, le taux de réoperculation est élevé dans toutes les colonies infestées (figures 3 et S1), atteignant les niveaux les plus élevés dans les colonies résistantes aux acariens. Partout au Brésil et en Afrique, la lutte organisée contre les parasites apicoles est minime et les pressions de sélection pour de tels caractères ont donc toujours été élevées. La gestion constante d‘un large éventail de ravageurs et de pathogènes du couvain dans l‘hémisphère Nord supprime une grande partie de cette pression sélective. Dans cette étude et dans des études antérieures, la capacité de détecter les acariens (figure S1) et de retirer le couvain infesté (tableau S3) est très variable. Il ne fait aucun doute que la composition des colonies joue un rôle puisque la réoperculation s‘est produite le plus dans les colonies mixtes plutôt que dans les colonies hautement hygiéniques ou hautement non hygiéniques.

Principales références
- Vidéos : You tube : Reinertsen honninggård
- 1.Oddie, M., Dahle, B., Neumann, P. (2017) Norwegian honey bees surviving Varroa destructormite infestations by means of natural selection. PeerJ 5, e3956. https://doi.org/10.7717/peerj.3956
- 2.Oddie, M. A. Y., Büchler, R., Dahle, B., Kovacic, M., Le Conte, Locke, B., de Miranda, J., Mondet, F., Neumann, P. (2018). Rapid parallel evolution overcomes global honey bee parasite. Sci. Rep. 8, 7704
- 3.Martin, S.J., Hawkins, G.P., Brettell, L.E. et al. Varroa destructor reproduction and cell re-capping in mite-resistant Apis mellifera populations. Apidologie (2019). https://doi.org/10.1007/s13592-019-00721-9
- 4.Erik Österlund 2020, 8,5 miljoner till forskning pa varroaresistenta bin – i Norge Gadden n°1/2020 4-7