Leur physiologie est particulièrement adaptée à cette fonction : l’appareil reproducteur occupe une bonne partie de l’abdomen du faux-bourdon. La performance est recherchée avant tout : gros yeux élaborés, récepteurs développés dans les antennes, puissance et rapidité en vol… L’accouplement en vol relève de la prouesse et il y a concurrence pour transmettre son patrimoine génétique dans la cohorte de mâles qui poursuit la reine lors du vol nuptial.
L’appareil génital du faux-bourdon est composé, d’une part, d’un ensemble comprenant les testicules, les glandes à mucus et les vésicules séminales organisées par paires. Cet ensemble est situé dans la zone antéro-dorsale de l’abdomen. D’autre part, l’endophalus, incluant le pénis, le bulbe et le canal éjaculateur, forme une poche repliée à l’intérieur de l’abdomen.
Les testicules produisent le sperme qui migre ensuite vers les vésicules séminales. Cette migration commence 2 jours avant ou 3 à 5 jours après l’émergence de l’individu selon les sources. C’est entre le 3° et le 7° jour du stade adulte que les vésicules séminales sont complètement remplies. Les testicules sont à leur maximum de développement (5- 6 mm de long) pendant la nymphose et se rétractent progressivement ensuite pour finir par devenir une paire d’appendices plats d’environ 1,5 mm de long. Les organes reproducteurs mâles sont complètement mûrs entre le 9° et le 12° jour mais la maturité sexuelle effective est plus tardive, prenant en considération d’autres éléments comme l’expérience du vol par exemple. Les auteurs envisagent la pleine maturité sexuelle en moyenne autour du 20° jour.
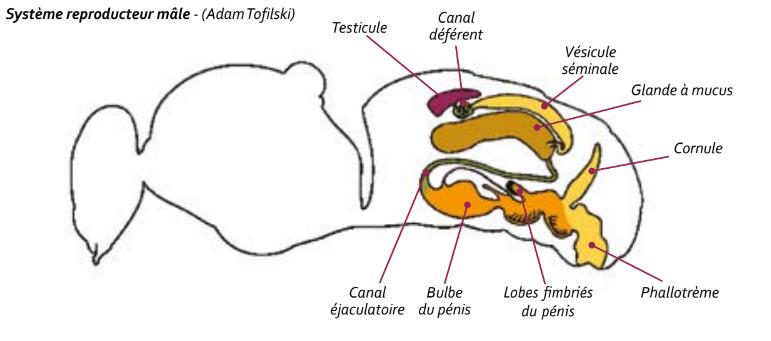
Les testicules et les vésicules séminales sont reliées par le canal déférent, un conduit courbe spiralé. Les vésicules séminales sont à l’opposé reliées aux glandes à mucus qui commencent leur sécrétion deux jours avant l’émergence avec un plein potentiel de développement au 5° jour du stade adulte, c’est-à
dire avant la maturité des vésicules séminales. Le mucus est de plus en plus visqueux au fur et à mesure du stade de maturation et donc de l’âge du faux-bourdon. Le mucus est impliqué dans la survie des spermatozoïdes, au même titre que les sécrétions glandulaires produites par la reine dans la spermathèque. Les profils protéiques du liquide séminal des mâles et du liquide spermathécal de la reine sont très différents, ce qui permet aux chercheurs de conclure que mâles et reines sont conjointement impliqués dans la fonction de viabilité des spermatozoïdes mais d’une manière différente.
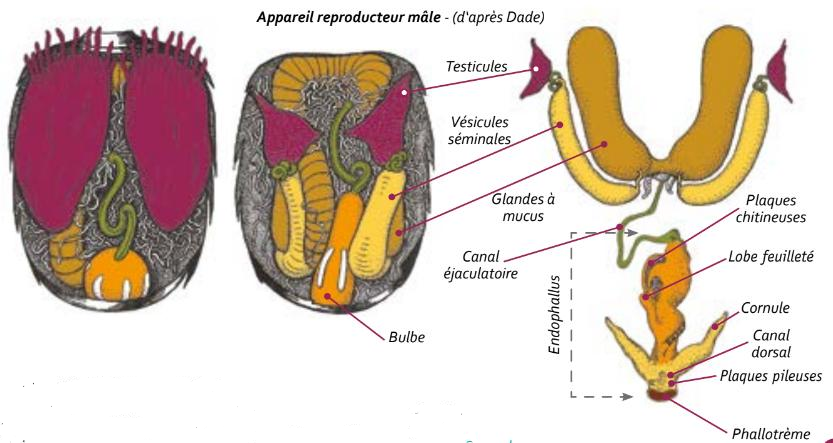
Par contraction musculaire, le sperme et le contenu des glandes à mucus passent par le canal éjaculatoire au moment de la copulation. Les vésicules séminales sont alors vidées, puis les glandes à mucus. L’épithélium des glandes à mucus est détruit et éjaculé également.
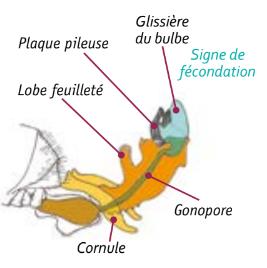
Au moment de l’éversion, l’endophalus se déploie vers l’extérieur : la paroi interne devient externe. En premier lieu, le bulbe et les plaques chitineuses sont poussés vers l’extérieur de l’abdomen. Sous une pression qui monte en intensité, bulbe et plaques chitineuses passent le conduit du cervix (col de l’utérus). L’éversion du bulbe se poursuit et, en phase finale, glandes à mucus et vésicules séminales se retrouvent dans une poussée hors de l’abdomen du mâle. Sperme et mucus sont transférés vers les oviductes de la reine qui conserve un « signe de fécondation » à l’issue de l’accouplement : les plaques chitineuses et la glissière du bulbe qui se sont séparées de l’endophallus du mâle désormais mort.
Quelques chiffres sur la copulation naturelle :
7 à 17 mâles (variations possibles) fécondent une seule reine lors d’un vol de fécondation.
Chaque copulation ne dure que quelques secondes.
15 à 30 % des reines meurent lors du vol de fécondation.
5,3 à 5,7 millions de spermatozoïdes peuvent entrer dans la spermathèque.
Au cours de sa vie, une reine pond entre 300.000 et 500.000 œufs.
Bibliographie
Dade, H. A.(1994). Anatomy and dissection of the honeybee. IBRA.
den Boer, S. P., Boomsma, J. J., & Baer, B. (2009). Honey bee males and queens use glandular secretions to enhance sperm viability before and after storage. Journal of Insect Physiology, 55(6), 538-543.
Moritz, R. F.(1985). The effects of multiple mating on the worker-queen conflict in Apis mellifera
L. Behavioral Ecology and Sociobiology, 16(4), 375-377.
Ruttner, F. (1968). Sexualité et reproduction. Chauvin, R. : Traité de Biologie d’Abeille. Masson et Cie. Paris, 1, 145-185.
Winston, M. L. (1991). The biology of the honey bee. Harvard University Press.
Woyke, J. (1955). Multiple mating of the honeybee queen (Apis mellifica L.) in one nuptial flight.
Bull. Acad. Polon. Sci. Cl, 3, 175-180.